

How Does A Bacterial Cell Know Which Strand To Replace When Carrying Out Mismatch Repair?
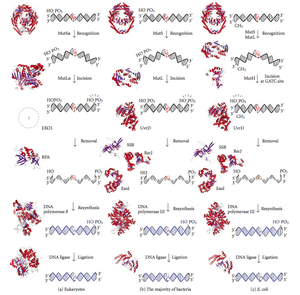
Diagram of Deoxyribonucleic acid mismatch repair pathways. The outset column depicts mismatch repair in eukaryotes, while the second depicts repair in almost bacteria. The third cavalcade shows mismatch repair, to be specific in Due east. coli.
DNA mismatch repair (MMR) is a organization for recognizing and repairing erroneous insertion, deletion, and mis-incorporation of bases that can arise during DNA replication and recombination, as well as repairing some forms of Dna damage.[1] [2]
Mismatch repair is strand-specific. During Deoxyribonucleic acid synthesis the newly synthesised (daughter) strand will commonly include errors. In order to brainstorm repair, the mismatch repair machinery distinguishes the newly synthesised strand from the template (parental). In gram-negative leaner, transient hemimethylation distinguishes the strands (the parental is methylated and daughter is not). However, in other prokaryotes and eukaryotes, the exact machinery is not clear. It is suspected that, in eukaryotes, newly synthesized lagging-strand Deoxyribonucleic acid transiently contains nicks (before beingness sealed by DNA ligase) and provides a signal that directs mismatch proofreading systems to the advisable strand. This implies that these nicks must exist nowadays in the leading strand, and bear witness for this has recently been institute.[3] Recent piece of work[4] has shown that nicks are sites for RFC-dependent loading of the replication sliding clamp, proliferating cell nuclear antigen (PCNA), in an orientation-specific fashion, such that one face of the donut-shape poly peptide is juxtaposed toward the three'-OH end at the nick. Loaded PCNA and then directs the activity of the MutLalpha endonuclease [v] to the girl strand in the presence of a mismatch and MutSalpha or MutSbeta.
Any mutational event that disrupts the superhelical construction of DNA carries with information technology the potential to compromise the genetic stability of a cell. The fact that the damage detection and repair systems are equally complex as the replication machinery itself highlights the importance evolution has attached to Dna fidelity.
Examples of mismatched bases include a G/T or A/C pairing (meet DNA repair). Mismatches are commonly due to tautomerization of bases during DNA replication. The damage is repaired by recognition of the deformity caused by the mismatch, determining the template and non-template strand, and excising the wrongly incorporated base of operations and replacing it with the right nucleotide. The removal process involves more than merely the mismatched nucleotide itself. A few or upwards to thousands of base pairs of the newly synthesized Deoxyribonucleic acid strand can be removed.
Mismatch repair proteins [edit]
| Dna mismatch repair poly peptide, C-terminal domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 hpms2-atpgs | |||||||||
| Identifiers | |||||||||
| Symbol | DNA_mis_repair | ||||||||
| Pfam | PF01119 | ||||||||
| Pfam clan | CL0329 | ||||||||
| InterPro | IPR013507 | ||||||||
| PROSITE | PDOC00057 | ||||||||
| SCOP2 | 1bkn / Telescopic / SUPFAM | ||||||||
| |||||||||
Mismatch repair is a highly conserved process from prokaryotes to eukaryotes. The commencement evidence for mismatch repair was obtained from South. pneumoniae (the hexA and hexB genes). Subsequent work on E. coli has identified a number of genes that, when mutationally inactivated, crusade hypermutable strains. The gene products are, therefore, called the "Mut" proteins, and are the major active components of the mismatch repair arrangement. Three of these proteins are essential in detecting the mismatch and directing repair machinery to it: MutS, MutH and MutL (MutS is a homologue of HexA and MutL of HexB).
MutS forms a dimer (MutS2) that recognises the mismatched base on the daughter strand and binds the mutated Dna. MutH binds at hemimethylated sites along the girl Deoxyribonucleic acid, but its action is latent, existence activated only upon contact by a MutL dimer (MutLii), which binds the MutS-Deoxyribonucleic acid circuitous and acts every bit a mediator between MutS2 and MutH, activating the latter. The Deoxyribonucleic acid is looped out to search for the nearest d(GATC) methylation site to the mismatch, which could exist upwardly to one kb away. Upon activation past the MutS-DNA complex, MutH nicks the girl strand near the hemimethylated site. MutL recruits UvrD helicase (DNA Helicase Two) to divide the two strands with a specific iii' to 5' polarity. The entire MutSHL complex then slides along the Dna in the direction of the mismatch, liberating the strand to be excised equally it goes. An exonuclease trails the complex and digests the ss-DNA tail. The exonuclease recruited is dependent on which side of the mismatch MutH incises the strand – 5' or three'. If the nick made by MutH is on the v' end of the mismatch, either RecJ or ExoVII (both 5' to 3' exonucleases) is used. If, however, the nick is on the three' end of the mismatch, ExoI (a iii' to five' enzyme) is used.
The entire process ends past the mismatch site - i.due east., both the site itself and its surrounding nucleotides are fully excised. The unmarried-strand gap created by the exonuclease tin can then be repaired by DNA Polymerase III (assisted by single-strand-binding protein), which uses the other strand as a template, and finally sealed by DNA ligase. Dna methylase so speedily methylates the daughter strand.
MutS homologs [edit]
When bound, the MutS2 dimer bends the Deoxyribonucleic acid helix and shields approximately 20 base pairs. It has weak ATPase activeness, and binding of ATP leads to the formation of 3rd structures on the surface of the molecule. The crystal structure of MutS reveals that it is exceptionally asymmetric, and, while its agile conformation is a dimer, but 1 of the 2 halves interacts with the mismatch site.
In eukaryotes, MutSouthward homologs course two major heterodimers: Msh2/Msh6 (MutSα) and Msh2/Msh3 (MutSβ). The MutSα pathway is involved primarily in base of operations commutation and small-loop mismatch repair. The MutSβ pathway is too involved in pocket-size-loop repair, in addition to big-loop (~10 nucleotide loops) repair. However, MutSβ does non repair base substitutions.
MutL homologs [edit]
MutL also has weak ATPase activity (it uses ATP for purposes of move). It forms a complex with MutS and MutH, increasing the MutS footprint on the Dna.
All the same, the processivity (the distance the enzyme can movement along the DNA before dissociating) of UvrD is just ~40–fifty bp. Considering the distance between the nick created by MutH and the mismatch can average ~600 bp, if there is non another UvrD loaded the unwound section is then gratuitous to re-anneal to its complementary strand, forcing the process to start over. Even so, when assisted by MutL, the rate of UvrD loading is greatly increased. While the processivity (and ATP utilisation) of the private UvrD molecules remains the aforementioned, the total event on the DNA is boosted considerably; the Deoxyribonucleic acid has no hazard to re-anneal, as each UvrD unwinds twoscore-50 bp of Dna, dissociates, then is immediately replaced past another UvrD, repeating the process. This exposes large sections of Deoxyribonucleic acid to exonuclease digestion, assuasive for quick excision (and later replacement) of the incorrect Deoxyribonucleic acid.
Eukaryotes take five GrandutL homologs designated equally MLH1, MLH2, MLH3, PMS1, and PMS2. They grade heterodimers that mimic MutL in E. coli. The human homologs of prokaryotic MutL class iii complexes referred to as MutLα, MutLβ, and MutLγ. The MutLα complex is fabricated of MLH1 and PMS2 subunits, the MutLβ heterodimer is made of MLH1 and PMS1, whereas MutLγ is fabricated of MLH1 and MLH3. MutLα acts as an endonuclease that introduces strand breaks in the daughter strand upon activation by mismatch and other required proteins, MutSα and PCNA. These strand interruptions serve as entry points for an exonuclease activity that removes mismatched Deoxyribonucleic acid. Roles played past MutLβ and MutLγ in mismatch repair are less-understood.
MutH: an endonuclease present in E. coli and Salmonella [edit]
MutH is a very weak endonuclease that is activated one time bound to MutL (which itself is bound to MutS). It nicks unmethylated DNA and the unmethylated strand of hemimethylated DNA but does not nick fully methylated Deoxyribonucleic acid. Experiments have shown that mismatch repair is random if neither strand is methylated.[ commendation needed ] These behaviours led to the proposal that MutH determines which strand contains the mismatch. MutH has no eukaryotic homolog. Its endonuclease function is taken upward past MutL homologs, which have some specialized 5'-three' exonuclease action. The strand bias for removing mismatches from the newly synthesized daughter strand in eukaryotes may be provided past the gratuitous 3' ends of Okazaki fragments in the new strand created during replication.
PCNA β-sliding clench [edit]
PCNA and the β-sliding clamp associate with MutSα/β and MutS, respectively. Although initial reports suggested that the PCNA-MutSα complex may enhance mismatch recognition,[6] it has been recently demonstrated[seven] that there is no apparent modify in analogousness of MutSα for a mismatch in the presence or absenteeism of PCNA. Furthermore, mutants of MutSα that are unable to interact with PCNA in vitro exhibit the chapters to carry out mismatch recognition and mismatch excision to virtually wild type levels. Such mutants are defective in the repair reaction directed past a 5' strand interruption, suggesting for the first time MutSα function in a post-excision footstep of the reaction.
Clinical significance [edit]
Inherited defects in mismatch repair [edit]
Mutations in the human being homologues of the Mut proteins affect genomic stability, which can effect in microsatellite instability (MSI), implicated in some human cancers. In specific, the hereditary nonpolyposis colorectal cancers (HNPCC or Lynch syndrome) are attributed to damaging germline variants in the genes encoding the MutS and MutL homologues MSH2 and MLH1 respectively, which are thus classified as neoplasm suppressor genes. I subtype of HNPCC, the Muir-Torre Syndrome (MTS), is associated with skin tumors. If both inherited copies (alleles) of a MMR cistron carry damaging genetic variants, this results in a very rare and severe status: the mismatch repair cancer syndrome (or constitutional mismatch repair deficiency, CMMR-D), manifesting equally multiple occurrences of tumors at an early age, often colon and brain tumors.[8]
Epigenetic silencing of mismatch repair genes [edit]
Sporadic cancers with a Deoxyribonucleic acid repair deficiency simply rarely have a mutation in a Deoxyribonucleic acid repair cistron, just they instead tend to take epigenetic alterations such equally promoter methylation that inhibit Deoxyribonucleic acid repair gene expression.[ix] Virtually 13% of colorectal cancers are deficient in DNA mismatch repair, commonly due to loss of MLH1 (9.viii%), or sometimes MSH2, MSH6 or PMS2 (all ≤one.5%).[10] For almost MLH1-scarce desultory colorectal cancers, the deficiency was due to MLH1 promoter methylation.[10] Other cancer types have higher frequencies of MLH1 loss (run into table beneath), which are once more largely a result of methylation of the promoter of the MLH1 gene. A different epigenetic mechanism underlying MMR deficiencies might involve over-expression of a microRNA, for example miR-155 levels inversely correlate with expression of MLH1 or MSH2 in colorectal cancer.[11]
| Cancer type | Frequency of deficiency in cancer | Frequency of deficiency in next field defect |
|---|---|---|
| Tum | 32%[12] [13] | 24%-28% |
| Stomach (foveolar blazon tumors) | 74%[14] | 71% |
| Stomach in loftier-incidence Kashmir Valley | 73%[15] | 20% |
| Esophageal | 73%[16] | 27% |
| Caput and neck squamous cell carcinoma (HNSCC) | 31%-33%[17] [eighteen] | 20%-25% |
| Non-small-scale cell lung cancer (NSCLC) | 69%[19] | 72% |
| Colorectal | ten%[10] |
MMR failures in field defects [edit]
A field defect (field cancerization) is an surface area of epithelium that has been preconditioned by epigenetic or genetic changes, predisposing information technology towards evolution of cancer. Every bit pointed out past Rubin " ...there is show that more than than 80% of the somatic mutations found in mutator phenotype human colorectal tumors occur before the onset of concluding clonal expansion."[20] [21] Similarly, Vogelstein et al.[22] signal out that more than half of somatic mutations identified in tumors occurred in a pre-neoplastic phase (in a field defect), during growth of manifestly normal cells.
MLH1 deficiencies were common in the field defects (histologically normal tissues) surrounding tumors; see Tabular array above. Epigenetically silenced or mutated MLH1 would probable not confer a selective advantage upon a stalk cell, however, it would crusade increased mutation rates, and ane or more of the mutated genes may provide the cell with a selective advantage. The scarceMLH1 gene could then be carried along as a selectively about-neutral passenger (hitch-hiker) cistron when the mutated stem cell generates an expanded clone. The continued presence of a clone with an epigenetically repressed MLH1 would continue to generate farther mutations, some of which could produce a tumor.
MMR components in humans [edit]
In humans, seven DNA mismatch repair (MMR) proteins (MLH1, MLH3, MSH2, MSH3, MSH6, PMS1 and PMS2) work coordinately in sequential steps to initiate repair of Deoxyribonucleic acid mismatches.[23] In addition, at that place are Exo1-dependent and Exo1-independent MMR subpathways.[24]
Other gene products involved in mismatch repair (subsequent to initiation past MMR genes) in humans include Deoxyribonucleic acid polymerase delta, PCNA, RPA, HMGB1, RFC and DNA ligase I, plus histone and chromatin modifying factors.[25] [26]
In certain circumstances, the MMR pathway may recruit an error-prone Deoxyribonucleic acid polymerase eta (POLH). This happens in B-lymphocytes during somatic hypermutation, where POLH is used to introduce genetic variation into antibody genes.[27] However, this error-prone MMR pathway may be triggered in other types of human being cells upon exposure to genotoxins [28] and indeed it is broadly active in various human cancers, causing mutations that bear a signature of POLH activeness.[29]
MMR and mutation frequency [edit]
Recognizing and repairing mismatches and indels is important for cells because failure to do so results in microsatellite instability (MSI) and an elevated spontaneous mutation rate (mutator phenotype). In comparing to other cancer types, MMR-deficient (MSI) cancer has a very high frequency of mutations, close to melanoma and lung cancer,[xxx] cancer types caused past much exposure to UV radiations and mutagenic chemicals.
In addition to a very high mutation brunt, MMR deficiencies result in an unusual distribution of somatic mutations across the man genome: this suggests that MMR preferentially protects the factor-rich, early-replicating euchromatic regions.[31] In dissimilarity, the cistron-poor, late-replicating heterochromatic genome regions exhibit high mutation rates in many human tumors.[32]
The histone modification H3K36me3, an epigenetic marker of agile chromatin, has the power to recruit the MSH2-MSH6 (hMutSα) complex.[33] Consistently, regions of the human genome with high levels of H3K36me3 accumulate less mutations due to MMR activeness.[29]
Loss of multiple DNA repair pathways in tumors [edit]
Lack of MMR often occurs in coordination with loss of other Dna repair genes.[nine] For instance, MMR genes MLH1 and MLH3 equally well equally xi other DNA repair genes (such equally MGMT and many NER pathway genes) were significantly down-regulated in lower course as well equally in higher grade astrocytomas, in contrast to normal brain tissue.[34] Moreover, MLH1 and MGMT expression was closely correlated in 135 specimens of gastric cancer and loss of MLH1 and MGMT appeared to be synchronously accelerated during tumor progression.[35]
Deficient expression of multiple DNA repair genes is frequently establish in cancers,[nine] and may contribute to the thousands of mutations ordinarily found in cancers (meet Mutation frequencies in cancers).
Aging [edit]
A popular thought, that has failed to gain significant experimental support, is the idea that mutation, as distinct from DNA damage, is the primary cause of aging. Mice defective in the mutL homolog Pms2 take about a 100-fold elevated mutation frequency in all tissues, but do not appear to age more rapidly.[36] These mice display by and large normal evolution and life, except for early onset carcinogenesis and male infertility.
See also [edit]
- Base excision repair
- Nucleotide excision repair
References [edit]
- ^ Iyer RR, Pluciennik A, Burdett Five, Modrich PL (February 2006). "DNA mismatch repair: functions and mechanisms". Chemical Reviews. 106 (2): 302–23. doi:10.1021/cr0404794. PMID 16464007.
- ^ Larrea AA, Lujan SA, Kunkel TA (May 2010). "SnapShot: Dna mismatch repair". Jail cell. 141 (4): 730–730.e1. doi:10.1016/j.cell.2010.05.002. PMID 20478261. S2CID 26969788.
- ^ Heller RC, Marians KJ (December 2006). "Replisome associates and the direct restart of stalled replication forks". Nature Reviews. Molecular Cell Biology. 7 (12): 932–43. doi:10.1038/nrm2058. PMID 17139333. S2CID 27666329.
- ^ Pluciennik A, Dzantiev Fifty, Iyer RR, Constantin N, Kadyrov FA, Modrich P (September 2010). "PCNA office in the activation and strand direction of MutLα endonuclease in mismatch repair". Proceedings of the National University of Sciences of the United States of America. 107 (37): 16066–71. doi:x.1073/pnas.1010662107. PMC2941292. PMID 20713735.
- ^ Kadyrov FA, Dzantiev L, Constantin N, Modrich P (July 2006). "Endonucleolytic function of MutLalpha in human mismatch repair". Prison cell. 126 (2): 297–308. doi:x.1016/j.jail cell.2006.05.039. PMID 16873062. S2CID 15643051.
- ^ Flores-Rozas H, Clark D, Kolodner RD (November 2000). "Proliferating cell nuclear antigen and Msh2p-Msh6p interact to form an active mispair recognition complex". Nature Genetics. 26 (iii): 375–8. doi:10.1038/81708. PMID 11062484. S2CID 20861705.
- ^ Iyer RR, Pohlhaus TJ, Chen Due south, Hura GL, Dzantiev L, Beese LS, Modrich P (May 2008). "The MutSalpha-proliferating cell nuclear antigen interaction in human being Dna mismatch repair". The Periodical of Biological Chemistry. 283 (19): 13310–9. doi:x.1074/jbc.M800606200. PMC2423938. PMID 18326858.
- ^ Online Mendelian Inheritance in Human (OMIM): 276300
- ^ a b c Bernstein C, Bernstein H (May 2022). "Epigenetic reduction of DNA repair in progression to gastrointestinal cancer". Earth Journal of Gastrointestinal Oncology. 7 (5): 30–46. doi:ten.4251/wjgo.v7.i5.30. PMC4434036. PMID 25987950.
- ^ a b c Truninger M, Menigatti M, Luz J, Russell A, Haider R, Gebbers JO, et al. (May 2005). "Immunohistochemical assay reveals high frequency of PMS2 defects in colorectal cancer". Gastroenterology. 128 (5): 1160–71. doi:10.1053/j.gastro.2005.01.056. PMID 15887099.
- ^ Valeri N, Gasparini P, Fabbri M, Braconi C, Veronese A, Lovat F, et al. (Apr 2010). "Modulation of mismatch repair and genomic stability by miR-155". Proceedings of the National Academy of Sciences of the Us of America. 107 (15): 6982–7. Bibcode:2010PNAS..107.6982V. doi:x.1073/pnas.1002472107. PMC2872463. PMID 20351277.
- ^ Kupčinskaitė-Noreikienė R, Skiecevičienė J, Jonaitis 50, Ugenskienė R, Kupčinskas J, Markelis R, et al. (2013). "CpG island methylation of the MLH1, MGMT, DAPK, and CASP8 genes in cancerous and adjacent noncancerous stomach tissues". Medicina. 49 (8): 361–6. doi:x.3390/medicina49080056. PMID 24509146.
- ^ Waki T, Tamura G, Tsuchiya T, Sato K, Nishizuka S, Motoyama T (Baronial 2002). "Promoter methylation status of E-cadherin, hMLH1, and p16 genes in nonneoplastic gastric epithelia". The American Periodical of Pathology. 161 (ii): 399–403. doi:ten.1016/S0002-9440(10)64195-eight. PMC1850716. PMID 12163364.
- ^ Endoh Y, Tamura One thousand, Ajioka Y, Watanabe H, Motoyama T (September 2000). "Frequent hypermethylation of the hMLH1 gene promoter in differentiated-type tumors of the stomach with the gastric foveolar phenotype". The American Journal of Pathology. 157 (iii): 717–22. doi:10.1016/S0002-9440(10)64584-1. PMC1949419. PMID 10980110.
- ^ Wani M, Afroze D, Makhdoomi Grand, Hamid I, Wani B, Bhat G, et al. (2012). "Promoter methylation status of Deoxyribonucleic acid repair gene (hMLH1) in gastric carcinoma patients of the Kashmir valley" (PDF). Asian Pacific Journal of Cancer Prevention. thirteen (eight): 4177–81. doi:10.7314/apjcp.2012.13.8.4177. PMID 23098428.
- ^ Chang Z, Zhang W, Chang Z, Song Thou, Qin Y, Chang F, et al. (January 2022). "Expression characteristics of FHIT, p53, BRCA2 and MLH1 in families with a history of oesophageal cancer in a region with a high incidence of oesophageal cancer". Oncology Messages. 9 (1): 430–436. doi:10.3892/ol.2014.2682. PMC4246613. PMID 25436004.
- ^ Tawfik HM, El-Maqsoud NM, Hak BH, El-Sherbiny YM (2011). "Head and neck squamous jail cell carcinoma: mismatch repair immunohistochemistry and promoter hypermethylation of hMLH1 gene". American Periodical of Otolaryngology. 32 (6): 528–36. doi:10.1016/j.amjoto.2010.11.005. PMID 21353335.
- ^ Zuo C, Zhang H, Spencer HJ, Vural Due east, Suen JY, Schichman SA, et al. (October 2009). "Increased microsatellite instability and epigenetic inactivation of the hMLH1 gene in head and neck squamous cell carcinoma". Otolaryngology–Head and Neck Surgery. 141 (4): 484–90. doi:10.1016/j.otohns.2009.07.007. PMID 19786217. S2CID 8357370.
- ^ Safar AM, Spencer H, Su X, Coffey One thousand, Cooney CA, Ratnasinghe LD, et al. (June 2005). "Methylation profiling of archived non-pocket-size cell lung cancer: a promising prognostic system". Clinical Cancer Inquiry. eleven (12): 4400–5. doi:10.1158/1078-0432.CCR-04-2378. PMID 15958624.
- ^ Rubin H (March 2022). "Fields and field cancerization: the preneoplastic origins of cancer: asymptomatic hyperplastic fields are precursors of neoplasia, and their progression to tumors can exist tracked past saturation density in civilisation". BioEssays. 33 (3): 224–31. doi:ten.1002/bies.201000067. PMID 21254148. S2CID 44981539.
- ^ Tsao JL, Yatabe Y, Salovaara R, Järvinen HJ, Mecklin JP, Aaltonen LA, et al. (February 2000). "Genetic reconstruction of private colorectal tumor histories". Proceedings of the National Academy of Sciences of the Usa. 97 (3): 1236–41. Bibcode:2000PNAS...97.1236T. doi:x.1073/pnas.97.iii.1236. PMC15581. PMID 10655514.
- ^ Vogelstein B, Papadopoulos Northward, Velculescu VE, Zhou S, Diaz LA, Kinzler KW (March 2022). "Cancer genome landscapes". Science. 339 (6127): 1546–58. Bibcode:2013Sci...339.1546V. doi:10.1126/science.1235122. PMC3749880. PMID 23539594.
- ^ Pal T, Permuth-Wey J, Sellers TA (August 2008). "A review of the clinical relevance of mismatch-repair deficiency in ovarian cancer". Cancer. 113 (4): 733–42. doi:x.1002/cncr.23601. PMC2644411. PMID 18543306.
- ^ Goellner EM, Putnam CD, Kolodner RD (August 2022). "Exonuclease ane-dependent and independent mismatch repair". Deoxyribonucleic acid Repair. 32: 24–32. doi:x.1016/j.dnarep.2015.04.010. PMC4522362. PMID 25956862.
- ^ Li GM (Jan 2008). "Mechanisms and functions of DNA mismatch repair". Cell Research. 18 (ane): 85–98. doi:ten.1038/cr.2007.115. PMID 18157157.
- ^ Li GM (July 2022). "New insights and challenges in mismatch repair: getting over the chromatin hurdle". DNA Repair. 19: 48–54. doi:10.1016/j.dnarep.2014.03.027. PMC4127414. PMID 24767944.
- ^ Chahwan R, Edelmann W, Scharff Doc, Roa S (August 2022). "AIDing antibody diversity by fault-prone mismatch repair". Seminars in Immunology. 24 (4): 293–300. doi:10.1016/j.smim.2012.05.005. PMC3422444. PMID 22703640.
- ^ Hsieh P (September 2022). "DNA mismatch repair: Dr. Jekyll and Mr. Hyde?". Molecular Cell. 47 (v): 665–half-dozen. doi:10.1016/j.molcel.2012.08.020. PMC3457060. PMID 22980456.
- ^ a b Supek F, Lehner B (July 2022). "Clustered Mutation Signatures Reveal that Error-Prone Dna Repair Targets Mutations to Active Genes". Cell. 170 (3): 534–547.e23. doi:10.1016/j.prison cell.2017.07.003. hdl:10230/35343. PMID 28753428.
- ^ Tuna Thousand, Amos CI (November 2022). "Genomic sequencing in cancer". Cancer Letters. 340 (2): 161–70. doi:x.1016/j.canlet.2012.11.004. PMC3622788. PMID 23178448.
- ^ Supek F, Lehner B (May 2022). "Differential DNA mismatch repair underlies mutation rate variation across the human genome". Nature. 521 (7550): 81–4. Bibcode:2015Natur.521...81S. doi:x.1038/nature14173. PMC4425546. PMID 25707793.
- ^ Schuster-Böckler B, Lehner B (Baronial 2022). "Chromatin organization is a major influence on regional mutation rates in human cancer cells". Nature. 488 (7412): 504–vii. Bibcode:2012Natur.488..504S. doi:10.1038/nature11273. PMID 22820252. S2CID 205229634.
- ^ Li F, Mao G, Tong D, Huang J, Gu L, Yang W, Li GM (Apr 2022). "The histone mark H3K36me3 regulates human Dna mismatch repair through its interaction with MutSα". Prison cell. 153 (3): 590–600. doi:ten.1016/j.cell.2013.03.025. PMC3641580. PMID 23622243.
- ^ Jiang Z, Hu J, Li X, Jiang Y, Zhou Due west, Lu D (December 2006). "Expression analyses of 27 Deoxyribonucleic acid repair genes in astrocytoma by TaqMan low-density assortment". Neuroscience Letters. 409 (two): 112–7. doi:x.1016/j.neulet.2006.09.038. PMID 17034947.
- ^ Kitajima Y, Miyazaki Grand, Matsukura S, Tanaka Chiliad, Sekiguchi M (2003). "Loss of expression of Dna repair enzymes MGMT, hMLH1, and hMSH2 during tumor progression in gastric cancer". Gastric Cancer. half-dozen (2): 86–95. doi:10.1007/s10120-003-0213-z. PMID 12861399.
- ^ Narayanan L, Fritzell JA, Bakery SM, Liskay RM, Glazer PM (Apr 1997). "Elevated levels of mutation in multiple tissues of mice deficient in the Deoxyribonucleic acid mismatch repair gene Pms2". Proceedings of the National Academy of Sciences of the United states of america. 94 (vii): 3122–seven. doi:10.1073/pnas.94.vii.3122. PMC20332. PMID 9096356.
Further reading [edit]
- Hsieh P, Yamane Grand (2008). "Dna mismatch repair: molecular mechanism, cancer, and ageing". Mechanisms of Ageing and Development. 129 (7–8): 391–407. doi:10.1016/j.mad.2008.02.012. PMC2574955. PMID 18406444.
- Iyer RR, Pluciennik A, Burdett V, Modrich PL (February 2006). "DNA mismatch repair: functions and mechanisms". Chemical Reviews. 106 (2): 302–23. doi:10.1021/cr0404794. PMID 16464007.
- Joseph N, Duppatla Five, Rao DN (2006). Prokaryotic DNA mismatch repair. Progress in Nucleic Acid Inquiry and Molecular Biological science. Vol. 81. pp. ane–49. doi:10.1016/S0079-6603(06)81001-9. ISBN9780125400817. PMID 16891168.
- Yang Westward (August 2000). "Structure and role of mismatch repair proteins". Mutation Research. 460 (3–4): 245–56. doi:10.1016/s0921-8777(00)00030-6. PMID 10946232.
- Griffiths JF, Gilbert WM, Lewontin RC, Wessler SR, Suzuki DT, Miller JH (2004). An introduction to genetic assay (eighth ed.). New York, NY: Freeman. ISBN978-0-7167-4939-4.
- Kunkel TA, Erie DA (2005). "DNA mismatch repair". Almanac Review of Biochemistry. 74: 681–710. doi:10.1146/annurev.biochem.74.082803.133243. PMID 15952900.
- Friedberg EC, Walker GC, Siede West, Wood RD, Schultz RA, Ellenberger (2005). DNA repair and mutagenesis (2nd ed.). Washington, D.C.: ASM Press. ISBN978-1-55581-319-ii.
External links [edit]
- DNA Repair
- DNA+Mismatch+Repair at the US National Library of Medicine Medical Discipline Headings (MeSH)

How Does A Bacterial Cell Know Which Strand To Replace When Carrying Out Mismatch Repair?,
Source: https://en.wikipedia.org/wiki/DNA_mismatch_repair
Posted by: molinafludersomand.blogspot.com
0 Response to "How Does A Bacterial Cell Know Which Strand To Replace When Carrying Out Mismatch Repair?"
Post a Comment